Մոնոսախարիդներ
Մոնոսախարիդներ կամ միաշաքարներ, մոնոզներ (հունարեն՝ monos - միակ, sacchar - շաքար), բազմատոմ սպիրտների ալդեհիդներ կամ կետոններ են, որոնց մոլեկուլը պարունակում է մեկ կարբոնիլային՝ ալդեհիդային կամ կետոնային[1], և մի քանի հիդրօքսիլային խմբեր։ Մոնոսախարիդները ըստ իրենց շղթայի մեջ գտնվող ածխածինների թվի լինում են տարբեր։ Նրանց անվանումն ունի «ոզա» վերջավորությունը, որն ավելացվում է ածխածինների թիվը ցույց տվող հուներան բառի վրա։

Մոնոսախարիդների ընդհանուր բանաձևն է CnH2nOn։ Բնության մեջ առավել տարածված են այն մոնոսախարիդները, որոնց մոլեկուլները պարունակում են 5 (պենտոզներ) կամ 6 (հեքսոզներ) ածխածնի ատոմ։ Ըստ մոլեկուլում առկա ածխածնի ատոմների թվի, միաշաքարները բաժանվում են հետևյալ դասերի.
- տրիոզներ (մոլեկուլը պարունակում է 3 ածխածնի ատոմ),
- տետրոզներ (չորս ածխածնի ատոմ),
- պենտոզներ (հինգ ածխածնի ատոմ),
- հեքսոզներ (վեց ածխածնի ատոմ) և այլն։
Բնության մեջ միաշաքարները հանդիպում են միայն պենտոզների։ Պենտոզներ են, օրինակ՝ ռիբոզը (C5H10O5) և դեզօքսիռիբոզը (C5H10O4)։ Դրանք մտնում են ՌՆԹ և ԴՆԹ կազմի մեջ և պայմանավորում են նուկլեինաթթուների անվան առաջին մասը։ Հեքսոզներին են պատկանում, օրինակ՝ գլյուկոզը, գալակտոզը, ֆրուկտոզը։
Մոնոսախարիդներ
խմբագրելՄոնոսախարիդները ըստ կառուցվածքի պոլիհիդրօքսիկարբոնիլային միացություններ են (պոլիհիդրօքսիալդեհիդներ և պոլիհիդրօքսիկետոններ)։ Նրանք պինդ, ջրում լավ լուծելի, հիմնականում քաղցրահամ, միացություններ են։ Ջրային լուծույթներն ունեն չեզոք ռեակցիա։
Մոնոսախարիդների դասակարգումը
խմբագրելՄոնոսախարիդների դասակարգումը հիմնված է կառուցվածքային յուրահատկությունների վրա։ Ըստ կարբոնիլային խմբի բնույթի տարբերում են ալդոզներ (-CHO) և կետոզներ (-C=O),իսկ շղթայում ածխածնի ատոմների թվի՝ տրիոզներ, տետրոզներ, պենտոզներ, հեքսոզներ և հեպտոզներ։
Կենսաբանական համակարգերում կարևոր նշանակություն ունեն պենտոզները և հեքսոզները, որոնք, հաշվի առնելով վերը նշված հատկանիշներից, բաժանվում են ալդոպենտոզների, ալդոհեքսոզների, կետոպենտոզների, կետոհեքսոզների։ Քանի որ յուրաքանչյուր մոնոսախարիդի կազմում կա մի քանի ասիմետրիկ ածխածնի ատոմ, հետևաբար մոնոսախարիդները առաջացնում են տարբեր ստերեոիզոմերներ (օրինակ, գլյուկոզ, մանոզ, գալակտոզ՝ բոլորը ալդոհեքսոզ են, մտնում են ալդոզների ընտանիքի մեջ և տարբերվում են իրենց ֆիզիկական հատկություններով)։

Մոնոսախարիդների ստերեոիզոմերների թիվը որոշվում է 2n բանաձևով, որտեղ n-ասիմետրիկ ածխածինների թիվն է։ Ալդոպենտոզների ստերեոիզոմերների թիվը հավասար է 8 (23),և նրանք կազմում են չորս զույգ էնանտիոմերներ։ Ալդոհեքսոզների ստերեոիզոմերների թիվը հավասար է 16 (24) է, նրանք կազմում են 8 զույգ էնանտիոմերներ։ Մոնոսախարիդները պատկանում են երկու ստերեոիզոմերների շարքի՝ D և L։ Ածխաջրերի պարզագույն ներկայացուցիչներն են ալդե- և կետոտրիոզները։

D և L շարքերի պատկանելիությունը որոշվում է ամենամեծ համարը կրող ասիմետրիկ ածխածնի կոնֆիգուրացիայով, այն համեմատելով D կամ L- գլիցերալդեհիդի կոնֆիգուրացիայի հետ։ Տվյալ մոնոշաքարի անվանումը որոշվում է բոլոր խիրալ կենտրոնների կառուցվածքով[2]։


Մոնոսախարիդները օպտիկապես ակտիվ միացություններ են.նրանց լուծույթները պտտում են բևեռացված լույսի հարթությունը։ Բևեռացված լույսի հարթության պտտման մեծությունը և ուղղությունը յուրահատուկ է յուրաքանչյուր ստերեոիզոմերի համար, կապված չէ D կամ L շարքերի պատկանելիության հետ և որոշվում է փորձնական ճանապարհով։ Բնական ծագում ունեցող մոնոսախարիդները հիմնականում D-շարքի ներկայացուցիչներն են[3]։
Մոնոսախարիդների կառուցվածքի ընդունված է պատկերել Ֆիշերի պրոյեկցիոն բանաձևերով, որոնցում ասիմետրիկ ածխածինը չի նշվում։
Մոնոսախարիդների ցիկլիկ կառուցվածքը
խմբագրել
Մոնոսախարիդները ցուցաբերում են բոլոր այն հատկությունները, որոնք պայմանավորված են նրանց կազմի մեջ մտնող ֆունկցիոնալ խմբերով։ Սակայն որոշ քիմիական հատկություններ հնարավոր չէ բացատրել ելնելով վերը բերված կառուցվածքից։
-
 β-D-ֆրուկտոֆուրանոզ
β-D-ֆրուկտոֆուրանոզ -
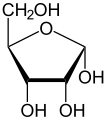 α-D-ռիբոֆուրանոզ
α-D-ռիբոֆուրանոզ -
 β-ռիբոֆուրանոզ
β-ռիբոֆուրանոզ -
 Պիրանոզ կառուցվածք Հեուորսի բանաձևով
Պիրանոզ կառուցվածք Հեուորսի բանաձևով




Առաջինը ռուս գիտնական Ա.Ա. Կոլինը կարծիք հայտնեց այն մասին, որ մոնոսախարիդներն ունեն ցիկլիկ կառուցվածք։ Լինելով հիդրօքսիալդեհիդներ (կամ կետոններ),մոնոսախարիդների բաց ձևերը ինքնաբերաբար կարող են վերածվել ցիկլիկ կառուցվածք ունեցող կիսաացետալների։ Օրինակ, ռիբոզի ճանկաձև (աքցանաձև) կոնֆիգուրացիայի կարբոնիլային խումբը և 4-րդ կամ 5-րդ ածխածնի OH խումբը տարածության մեջ մոտ են գտնվում և OH-ի թթվածնի նուկլեաֆիլ ատոմը գրոհում է կարբոնիլային ածխածնի էլեկտրաֆիլ ատոմի վրա։ Առաջանում է ցիկլիկ կիսաացետալ։ Կախված ցիկլիկ չափսերից, առաջացող միացությունները կոչվում են ֆուրանոզային (հինգանդամանի) կամ պիրանոզային (վեցանդամանի)։
Ցիկլիկ ձևում պրոխիրալ (կարբոնիլային խմբի ածխածինը) ոչ սիմետրիկ կենտրոնը վերածվում է խիրալ կենտրոնի։ Հետևաբար աճում է ստերեոիզոմերների թիվը։ Առաջացած կիսաացետալային OH խումբը կոչվում է գլիկոզիդային, իսկ ՛՛C-1՛՛ ածխածինը՝ անոմերային։ Ըստ գլիկոզիդային խմբի դիրքի տարբերում են α- և β-անոմերներ։ Ներմոլեկուլային փոխազդեցության հետևանքով առաջանում են 5 և 6 անդամանի թերմոդինամիկորեն կայուն ցիկլեր՝ ֆուրանոզային և պիրանոզային։
Մոնոսախարիդների ցիկլիկ կառուցվածքները ընդունված է պատկերել նաև Հեուորսի բանաձևերով։
Հեուորսի բանաձևերում պիրանոզային ցիկլի թթվածինը պատկերում են աջ հեռավոր անկյունում (C-1-ը՝ աջից)։OH-խմները, որոնք բաց ձևերի մոտ (Ֆիշերի բանաձևերում) գտնվում էին աջ կողմից գրվում են ցիկլի հարթությունից ցածր, իսկ ձախ կողմից գտնվող OH-խմբերը՝ հարթությունից վերև։ Այն անոմերները, որոնցում գլիկոզիդային խումբը պատկերում են հարթության տակ, կոչվում են α-անոմերներ։ Անոմերները, որոնցում գլիկոզիդային խումբը պատկերում են հարթությունից վեր կոչվում են β-անոմերներ։
Ցիկլո-օքսո տաուտոմերիա
խմբագրելԱնոմերները տարբերվում են հալման, եռման ջերմաստիճաններով, օպտիկական հատկություններով։ Հայտնի է, որ α-D-գլյուկոպիրանոզի թարմ լուծույթի տեսակարար պտտման անկյունը հավասար է 112°,իսկ β-D-գլյուկոպիրանոզինը՝ 19°։
Մոնոսախարիդի բոլոր 5 ձևերը (ացիկլիկ,α- և β-ֆուրանոզային և α- և β պիրանոզային) լուծույթում գտնվում են դինամիկ հավասարակշռության մեջ։ Առավել կայուն են մոնոսախարիդների պիրանոզային ձևերը։
Օրինակ, գլյուկոզի ջրային լուծույթում 99,9% կազմում է գլյուկոզի պիրանոզային ձևը։ Բաց ձևը կազմում է ընդամենը 0,003-0,03% և միջանկյալ է՝ ցիկլիկ ձևերը փոխարկվում են մեկը մյուսին բաց ձևի միջոցով։ Այսինքն գոյություն ունեն տաուտոմերիայի ևս մեկ տեսակ՝ ցիկլո-օքսո։
Դրանով է բացատրվում այն փաստը, որ բյուրեղային α- և β-D-գլյուկոպիրանոզներից պատրաստված թարմ լուծույթների բևեռացված լույսի պտտման անկյունը, որը համապատասխանաբար հավասար է +112° և +19°,մի քանի ժամ հետո փոխվում է և հավասարվում +52,7°։ Դա պայմանավորված է նրանով, որ լուծույթում հաստատվում է դինամիկ հավասարակշռություն տարբեր իզոմերների միջև, որում գերակշռում են α- և β-պիրանոզային ձևերը (1։2 հարաբերությամբ,36% α- և 64% β-)։ Մոնոսախարիդների թարմ լուծույթների ժամանակի ընթացքում բևեռացված լույսի պտտման անկյան փոփոխության երևույթը կոչվում է մուտառոտացիա։ Այդ պատճառով լուծույթում գտնվող մոնոսախարիդի համար սովորաբար չեն նշվում անոմերային ածխածնի կոնֆիգուրացիան։
-
β-D-ֆրուկտոֆուրանոզ
-
 α-D-ֆրուկտոֆուրանոզ
α-D-ֆրուկտոֆուրանոզ

Մոնոսախարիդների կոնֆորմացիաները
խմբագրելՔանի որ վեցանդամանի (պիրանոզային) ցիկլը հարթ չէ, Հեուորսի բանաձևերը լրիվ պատկերացում չեն տալիս մոնոսախարիդների ցիկլիկ ձևի տարածական կառուցվածքի մասին։ Ինչպես և ցիկլոհեքսանի դեպքում, ածխաջրերի առավել կայուն կոնֆորմացիան բազկաթոռն է։
-
 α-D-գլյուկոպիրանոզ (բազկաթոռ ձև)
α-D-գլյուկոպիրանոզ (բազկաթոռ ձև) -



Կոնֆորմացիոն կառուցվածքով է բացատրվում D-գլյուկոզի առավել կայուն լինելը՝ նրանում առաջնային սպիրտային և բոլոր հիդրօքսիլ խմբերը (բացառությամբ կիսաացետալային խմբի, այն էլ միայն α-անոմերի դեպքում) գտնվում են էկվատորիալ դիրքում։ Յուրաքանչյուր այլ հեքսոզ ունի գոնե մեկ աքսիալ OH-խումբ։
Կոնֆորմացիոն կառուցվածքների հարաբերական կայունությամբ է բացատրվում գլյուկոզի α- և β-պիրանոզային ձևերի քանակները լուծույթում։ Տաուտոմեր հավասարակշռությունը շեղված է դեպի β-D-անոմերի առաջացում, քանի որ առավել կայուն են տեղակալիչների էկվատորիալ դասավորություն ունեցող կոնֆորմերները։

Մոնոսախարիդի կոնֆիգուրացիան, հատկապես անոմերային ածխածնինը (α- կամ β-) չափազանց կարևոր է պոլիսախարիդների տարածական կառուցվածքի ձևավորման համար, էկվատորիալ դիրքերի դեպքում առաջացած շղթան ունի գծային կառուցվածք (ցելյուլոզ),իկս աքսիալ դասավորվածության դեպքում առաջանում են գալարներ և ձևավորվում է պարուրաձև երկրորդային կառուցվածք (օսլա)։
Իզոմերիա
խմբագրել
Սկսած տրիոզներից, մոնոսախարիդներն օպտիկապես ակտիվ են, այսինքն՝ պտտում են լույսի բևեռացած ճառագայթի տատանման հարթությունը։ Նյութերի օպտիկական ակտիվությունը կապված է ասիմետրիկ ածխածինների առկայության հետ։ Ասիմետրիկ է կոչվում այն ածխածինը, որի 4 ասիմետրիկ ածխածին, իսկ կետահեքսոզներն ու պենտոզները՝ 3։ Ըստ Վանտ Հոֆի բանաձևի ալդոհեքսոզների իզոմերների քանակը հավասար է 24=16-ի, իսկ կետոհեքսոզների և պենտոզների իզոմերների քանակը՝ 23=8-ի։
Կախված H և OH խմբերի տարածական դասավորությունից՝ C6H12O6 ալդոհեքսոզները հանդես են գալիս ութ իզոմերների ձևով (գլյուկոզ, մաննոզ, գալակտոզ, ալոզ, ալտրոզ, գուլոզ, իդոզ, տալոզ)։ Դրանցից յուրաքանչյուրն առաջացնում է 4 իզոմեր՝ α- և β- ձևերը ներառյալ, և իզոմերների թիվն այսպիսով հասնում է 32–ի։ Յուրաքանչյուր մոնոսախարիդ, սկասած տրիոզներից, ունի իր ստերիոիզոմերները, սյսինքն՝ հարթության վրա իր կառուցվածքի հայելանման պատկերին համապատասխանող նյութը։
Տեսակներ
խմբագրելՄոնոսախարիդների համարյա բոլոր ներկայացուցիչները անջատվել կամ սինթեզվել են, այնուամենայինիվ բնության մեջ գտնվում են նրանցից մի քանիսը միայն, ալդոհեքսոզներից՝ D-գլյուկոզան, D-գալակտոզան և D-մանոզան, իսկ կետոհեքսոզներից՝ միայն D-ֆրուկտոզան։ Պենտոզներից հայտնի են L-արաբինոզան, D-քսիլոզան, D-ռիբոզան և այլն։
Դասակարգում
խմբագրելԲնության մեջ հայտնաբերվել են հետևյալ մոնոզները, որոշ տեսակներ սինթեզվել են.
| Ալդոտրիոզներ |  գլիցերալդեհիդ (գլիցերալ) | |||||||
| Ալդոտետրոզներ |  D-էրիթրոզա |
 D-տրեոզա | ||||||
| Ալդոպենտոզներ |  D-ռիբոզա |
 D-արաբինոզա |
 D-քսիլոզա |
 D-լիկսոզա | ||||
| Ալդոհեքսոզներ |  D-ալլոզա |
 D-ալտրոզա |
 D-գլյուկոզա |
 D-մաննոզա |
 D-գուլոզա |
 D-իդոզա |
 D-գալակտոզա |
 D-տալոզա |
| Կետոտրիոզներ | Դիհիդրոքսիացետոն | |||
| կետոտետրոզներ |  էրիտրուլոզա | |||
| կետոպենտոզներ |  ռիբուլոզա |
 քսիլուլոզա | ||
| կետոհեքսոզներ |  պսիկոզա |
 ֆրուկտոզա |
 սորբոզա |
 տագատոզա |
Հայտնի են նաև մոնոսախարիդներ ավելի երկար շղթաներով, օրինակ՝ կետոհեքսոզներ, մաննոհեպտուլոզա և սեդոհեպտուլոզա։
Ալդոհեպտոզներ (D-շարք)։
D-գլիցերո-D-ալլոհեպտոզա
D-գլիցերո-D-ալտրոհեպտոզա
D-գլիցերո-D-գլյուկոհեպտոզա
D-գլիցերո-D-մաննոհեպտոզա
D-գլիցերո-D-գուլյոհեպտոզա
D-գլիցերո-D-իդոհեպտոզա
D-գլիցերո-D-գալակտոհեպտոզա
D-գլիցերո-D-տալոհեպտոզա
D-գլիցերո-L-ալլոհեպտոզա
D-գլիցերո-L-ալտրոհեպտոզա
D-գլիցերո-L-գլյուկոհեպտոզա
D-գլիցերո-L-մաննոհեպտոզա
D-գլիցերո-L-գուլոհեպտոզա
D-գլիցերո-L-իդոհեպտոզա
D-գլիցերո-L-գալակտոհեպտոզա
D-գլիցերո-L-տալոհեպտոզա
Քիմիական հատկություններ
խմբագրելՄոնոսախարիդների կարևոր հատկությունը դրանց ֆերմենտային խմորումն է՝ միկրոօրգանիզմների արտադրած օրգանական կատալիզատորների՝ ֆերմենտների ազդեցությամբ մոլեկուլի տրոհումը։ Խմորման են ենթարկվում հիմնականում հեքսոզները, և, կախված ազդող ֆերմենտների բնույթից, ընթանում են տարբեր ռեակցիաներ.
- սպիրտային խմորում. մեխանիզմը բարդ է, և արտահայտվում է հետևյալ հավասարումով.

-
- կաթնաթթու (α-հիդրօքսիպրոպիոնաթթու)

Մոնոսախարիդները հեշտությամբ օքսիդանում են, նրանք վերականգնում են արծաթի օքսիդի ամոնիակային լուծույթը և ֆելինգի հեղուկը։ Ալդոզների զգույշ օքսիդացման ժամանակ առաջանում են նույն թվով ածխածնային ատոմներ ունեցող միահիմն օքսիթթուներ, այսպես կոչվող օննաթթուներ։
Մոնոսախարիդները վերականգնելիս նրանք վերածվում են բազմատոմ սպիրտների։
Մոնոսախարիդների քիմիական հատկությունները
խմբագրելՄոնոսախարիդները, շնորհիվ հետերոֆունկցիոնալ կառուցվածքի (բազմաթիվ ռեակցիոն կենտրոնների առկայության) չափազանց ռեակցիոնունակ են։ Կիսաացետալային հիդրօքսիլով են պայմանավորված մոնոսախարիդների կարևորագույն հատկությունները։ Մոնոսախարիդները չոր HCl-ի ներկայությամբ սպիրտների հետ փոխազդելիս (անջուր պայմաններում),առաջացնում են ացետալներ, որոնք կոչվում են գլիկոզիդներ՝ կիսաացետալային խումբը փոխարինվում է նուկլեաֆիլ տեղակալման մեխանիզմով։ Առաջանում է α- և β- գլիկոզիդների խառնուրդ։

Անոմերային գլիկոզիդների խառնուրդը կարելի է պատկերել մեկ բանաձևով։
Մոնոսախարիդները գլիկոզիդներ են առաջացնում նաև մոնոսախարիդների սպիրտային խմբերի հաշվին, այսինքն առաջանում են օլիգո- և պոլիսախարիդներ։ Բացի այդ, օրգանիզմում տեղի է ունենում D-գլյուկուրոնաթթվի գլիկոզիդների առաջացում՝ որոշ տոքսիկ ֆենոլային միացությունների հետ։
Այդ գլիկոզիդները (գլյուկուրոնիդները) հեռանում են մեզով և տեղի է ունենում դետոքիկացում։O-գլիկոզիդները նման են ազոտ պարունակող անալոգները՝ N-գլիկոզիդները, որոնք հանդես են գալիս որպես նուկլեոտիդների, նուկլեինաթթուների բաղադրամասեր։

Օրգանիզմում գլիկոզիդների առաջացմանը մասնակցում են մոնոսախարիդների ֆոսֆատները կամ նուկլեոզիդդիֆոսֆատները։ Այս միացություններում ֆոսֆորական թթվի մնացորդը և նուկլեոզիդդիֆոսֆատը լավ հեռացող խմբեր են։
Գլիկոզիդները կարող են հիդրոլիզվել թթվային միջավայրում (ացետալների նման),բայց բավականին կայուն են նոսր հիմքերի հանդեպ։ Գլիկոզիդների հիդրոլիզը, ըստ էության, առաջացման ռեակցիայի հակառակն է՝ արդյունքում առաջանում է սպիրտ և մոնոսախարիդ։ Օրինակ, մեթիլ-β-D-գլյուկոպիրանոզիդի հիդրոլիզի արդյունքում առաջանում են գլյուկոզ և մեթանոլ։
Սպիրտային OH-խմբերի մասնակցությամբ ընթացող ռեակցիաներ.պարզ և բարդ եթերների առաջացում։
Պարզ եթերներ առաջանում են հիմնային միջավայրում գլիկոզիդների (կամ մոնոսախարիդների) և ալկիլհալոգենիդների (կամ այլ ալկիլացնող ագենտների) փոխազդեցության արդյունքում առաջացած գլիկոզիդների եթերների թթվային հիդրոլիզով։ Օրինակ, մեթիլ-α-D-գլյուկոպիրանոզի ալկիլացման արդյունքում առաջանում է մեթիլտետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզիդ։
Առաջացած մեթիլտետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզիդը ենթարկվում է թթվային հիդրոլիզի, վերածվելով տետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզի՝ եթերի։ Մոնոսախարիդների ալկիլացման ընթացքում նույնպես առաջանում են գլիկոզիդների եթերներ, որոնց գլիկոզիդային կապը թթվային միջավայրում ենթարկվում է հիդրոլիզի և առացանում է պարզ եթեր։
Բարդ եթերները առաջանում են մոնոսախարիդների և ացիլացնող ագենտների, օրինակ՝ կարբոնաթթուների անհիդրիդների, փոխազդեցության դեպքում։
Մոնոսախարիդների փոխարկումներում մեծ դեր ունեն ֆոսֆատները (ֆոսֆորական թթվի բարդ եթերները)։ Բացի արդեն հիշատակված գլյուկոզո-1-ֆոսֆատից, կարևոր նշանակություն ունեն գլյուկոզո-6-ֆոսֆատը, ֆրուկտոզո-6-ֆոսֆատը, դիֆոսֆատը, օրինակ՝ ֆրուկտոզ-1,6-դիֆոսֆատը (ֆրուկտոզ-1,6-բիֆոսֆատ)։
Մոնոսախարիդների էսթերները, ինչպես և բոլոր բարդ եթերները, ընդունակ են հիդրոլիզվելու թթվային և հիմնային միջավայրերում մինչև համապատասխան թթու (աղ՝ հիմնայինի դեպքում) և սպիրտ (տվյալ դեպքում մոնոսախարիդը սպիրտ է)։
Մոնոսախարիդների օքսիդացման ռեակցիաները։ Ալդոզների օքսիդացումը չեզոք և թթվային միջավայրերում ընթանում է առանց մոլեկուլների ածխածնային շղթայի փոփոխության։ Կախված օքսիդացման պայմաններից, կարող են օքսիդանալ միայն ալդեհիդային խումբը, ալդհեհիդային և սպիրտային խմբերը և այլն, և արդյունքում առաջանում են տարբեր միացություններ։
Օրինակ, գլյուկոզի օքսիդացման ժամանակ առաջանում են գլյուկոնաթթու, գլյուկուրոնաթթու, գլյուկարաթթու։
Մեղմ օքսիդիչի դեպքում (բրոմաջուր) օքսիդանում է միայն ալդեհիդային խումբը և ռեակցիայի արդյունքում առաջանում են ալդոնաթթուների, գլյուկոզի դեպքում՝ գլյուկոնաթթու։
Կենդանի օրգանիզմում նման օքսիդացումը ընթանում է համապատասխան ֆերմենտի մասնակցությամբ։
Սպիրտները, ինչպես հայտնի է, օքսիդանում են առաջացնելով կարբոնաթթուներ, ըստ որում առաջնային սպիրները երկրորդայինների հետ համեմատ օքսիդանում են ավելի արագ։
Ուժեղ օքսիդիչի ազդեցության ժամանակ (նոսր ազոտական թթու) օքսիդանում են և՛ ալդեհիդային, և՛ առաջնային սպիրտային խմբերը, առաջացնելով գլիկարաթթուներ կամ շաքարաթթուներ։
Այսպես, գլյուկոզի օքսիդացումից առաջանում է գլյուկարաթթու, մանոզի-մանարաթթու, գալակտոզի-գալակտարաթթու։
Կենդանի օրգանիզմներում կարևոր նշանակություն ունեն գլյուկուրոնաթթուները, որոնք առաջանում են առաջնային սպիրտային խմբի օքսիդացումից։ Օրինակ, գլյուկուրոնաթթուն, որը բազմազան ֆունկցիաներ է կատարում, մասնակցում է թունազրկման ռեակցիաներին, որպես կառուցվածքային հատված մտնում է պոլիսախարիդների կազմի մեջ և այլն։
Մոնոսախարիդներից ուրոնաթթուների առաջացման դեպքում անհրաժեշտ է «պաշտպանել» ալդեհիդային խումբը.օրինակ, օքսիդացումից առաջ, մոնոսախարիդը վերածել գլիկոզիդի։ Մոնոսախարիդների գլիկոզիդների օքսիդացման արդյունքում առաջանում են գլիկուրոնաթթուների գլիկոզիդներ, որոնց թթվային հիդրոլիզից առաջանում են ազատ գլիկուրոնաթթուներ։
Օրգանիզմում այդ պաշտպանական դերը կատարում է 1-ֆոսֆատային խումբը։
Մոնոսախարիդների վերականգնման հետևանքով առաջանում են ալդիտներ (շաքարային սպիրտներ),որոնք քաղցրահամ են։ Վերականգնումը ընթանում է խիստ պայմաններում, ուժեղ վերականգնիչների մասնակցությամբ։

Ալդիտներից մի քանիսը, օրինակ, քսիլիտի (D-քսիլոզից) և սորբիտը (սորբիտոլ,D-գլյուկոզից) շաքարախտով հիվանդները օգտագործում են որպես շաքարի փոխարինիչ։ Ալդիտները չեն ընդգրկվում նյութափոխանակության մեջ, համապատասխան ֆերմենտների բացակայության պատճառով։
Ալդոզների և կետոզների փոխադարձ փոխարկումը։ Հիմնային միջավայրում տեղի է ունենում մոնոսախարիդների իզոմերացում։CH-կենտրոնի առկայության շնորհիվ, հիմնային միջավայրում, տեղի է ունենում ալդոզներից կետոզների առաջացում (իզոմերացում) և էպիմերացում C-2 մոտ։
Օրգանիզմում նման փոխարկումները ընթանում են ֆերմենտների մասնակցությամբ։ Այս փոխարկումների նշանակությունը չափազանց մեծ է, քանի որ մոնոսախարիդներից կենդանական համակարգերում միայն գլյուկոզն ունի փոխանակման համար անհրաժեշտ ֆերմենտների համակարգ։ Հետևաբար, այլ մոնոշաքարները փոխանակվելու համար պիտի վերածվեն գլյուկոզի։ Նկարագրված եղանակով ֆրուկտոզից և մանոզից առաջանում է գլյուկոզ։
Մոնոսախարիդների հայտնաբերման ռեակցիաները
խմբագրելՊենտոզների և հեքսոզների տարբերիչ ռեակցիաները
խմբագրելՀանքային թթուների հետ տաքացնելիս մոնոսախարիդները ենթարկվում են դեհիդրատացման (կորցնում են երեք մոլեկուլ ջուր) և ալդոպենտոզներից առաջանում է ֆուրֆուրոլ, իսկ ալդո- և կետոհեքսոզներից՝ 5-հիդրօքսիմեթիլֆուրֆուրոլ։ Ֆուրֆուրոլը և 5-հիդրօքսիմեթիլֆուրֆուրոլը ֆենոլների և արոմատիկ ամինների հետ տալիս են կոնդեսացման ռեակցիաներ, առաջացնելով գունավոր միացություններ։ Այս ռեակցիաների վրա են հիմնված պենտոզների և հեքսոզների քանակական և որակական հայտնաբերման մի շարք եղանակներ։

Օրինակ, ֆուրֆուրոլը փոխազդելով անիլինի, իսկ 5-հիդրօքսիմեթիլֆուրֆուրոլը՝ ռեզորցինի հետ առաջացնում են կարմիր գույնի միացություններ։
Մոնոսախարիդների հայտնաբերման որակական ռեակցիաները
խմբագրելԱլդեհիդային (կարբոնիլային) խմբի օքսիդացման վրա են հիմնված մոնոսախարիդների հայտնաբերման (որակական) ռեակցիաները։ Չնայած ալդեհիդային (բաց) ձևի բաժինը տաուտոմեր խառնուրդում չափազանց փոքր է, բայց նա անընդհատ առաջանում է և մնում հաստատուն։ Տոլենսի (Ag+),Բենեդիկտի, Ֆելինգի (Cu2+) ռեակտիվների օգնությամբ կարելի է հայտնաբերել ալդոզները։ Հիմնային միջավայրում մետաղների իոնները վերականգնվում են, իսկ մոնոսախարիդները քայքայվում են՝ առաջացնելով օքսիդացման արդյունքների խառնուրդ։
RCHO+Ag(NH3)2+→Ag↓+օքսիդացման արդյունքներ
RCHO+Cu2+(կոմպլեքս)→Cu2O↓+օքսիդացման արդյունքներ
Այս ռեակտիվների հետ դրական ռեակցիա տվող մոնոսախարիդները և նրանց ածանցյալները կոչվում են վերականգնող շաքարներ։ Գլիկոզիդները չեն ցուցաբերում վերականգնիչ հատկություններ և չեն փոխազդում այս ռեագենտների հետ։ Սակայն, այս ռեակցիաները, ի տարբերությունյ բրոմաջրով և ազոտական թթվով օքսիդացման, չի կարելի օգտագործել ալդոնաթթուների ստացման համար, քանի որ հիմնային միջավայրում առաջանում է ենդիոլային ֆրագմենտ, և կարող է տեղի է ունենալ իզոմերացում (օրինակ գլյուկոզը վերածվի մանոզի կամ ֆրուկտոզի),ինչպես նաև ածխածնային շղթայի քայքայում։
Մոնոսախարիդների ածանցյալները
խմբագրելՄոնոսախարիդների ածանցյալներ են համարվում մոնոսախարիդային բնույթի այն միացությունները, որոնք մեկ կամ մի քանի հիդրօքսիլային խմբերի փոխարեն պարունակում են ջրածնի ատոմ կամ ֆունկցիոնալ խմբեր։ Օրինակ, նուկլեինաթթուների կազմի մեջ մտնում է 2-դեզօքսի-D-ռիբոզը, հետերոպոլիսախարիդների կազմի մեջ հանդիպում են մոնոսախարիդների ամինաածանցյալները։
Առավել կարևոր նշանակություն ունեն մոնոսախարիդների 2-ամինաածանցյալները, օրինակ,D-գլյուկոզամինը։

Բացի այդ, հանդիպում են նաև ամինախմբի ալկիլացված կամ ացիլացված ածանցյալներ՝ N-ացետիլգլյուկոզամին,N-ացետիլգալակտոզամին և այլն։

Նեյրամինաթթուն և նրա ածանցյալները, սիալաթթուները (N-ացետիլնեյրամինաթթուները) արյան և հյուսվածքների սպեցիֆիկ միացությունների բաղադրամասերն են։ Ազատ վիճակում գտնվում են ողնուղեղային հեղուկում։ Նեյրամինաթթվի սինթեզը կատարվում է պիրոխաղողաթթվից և մանոզամինից։


Ասկորբինաթթուն (վիտամին C) ըստ կառուցվածքի 2-կետո-L-գուլոնաթթվի γ-լակտոն է։

Ասկորբինաթթուն նյութափոխանակության ընթացքում մասնակցում է զանազան վերականգնման պրոցեսների։ Մասնավորապես կարևոր նշանակություն ունի կոլագենի կազմում պրոլինի հիդրօքսիլացմանը մասնակից ֆերմենտի (պրոլինհիդրօքսիլազի) գործունեության համար։ Ռեակցիայի արդյունքում ասկորբինաթթուն օքսիդանում է, վերածվելով դեհիդրոասկորբինաթթվի։

Արդյունաբերական եղանակներով ասկորբինաթթուն մեծ քանակներով ստանում են գլյուկոզից։ Վիտամին C-ի անբավարարության դեպքում զարգանում են զանազան հիվանդություններ (սկորբունտ, լնդախտ)։
Ստացում
խմբագրելՇաքարանյութի առաջին սինթեզը իրագործել է Ա. Մ. Բուտլերովը՝ 1861 թվականին։ Մրջնալդեհիդը տաքացնելով կրակաթի հետ, նա ստացավ բաց դեղնավուն քաղցր օշարակ, որն անվանեց մեթիլենիտան, մանիտանին (մաննիտի հին անունը) նման։ Այս օշարականման հեղուկը տալիս է մոնոսախարիդների սովորական ռեակցիաները։ Հետագայում նրանցից անջատվեց α-ակրոզան, այսինքն՝ ռեցեմիկ ֆրուկտոզան։
Որպես ինքնուրույն նյութ α-ակրոզան սինթեզվել է 1887 թվականին Է. Ֆիշերի կողմից՝ ակրոլեինից CH2=CH-CHO.ակրոլեինին բրոմ միացնելով ստացվել է դիբրոմիդ. որին վրա էլ բարիտաջուր ավելացնելիս տեղի է ունենում գլիցերինալդեհիդի առաջացում, որը մասամբ իզոմերելով դիօքսիացետոնի, փոխարկվում է α-ակրոզայի՝ Պարզվեց որ Է. Ֆիշերի ստացաշ ակրոզան բազմաթիվ շաքարների խառնուրդ է. նրա մեջ հայտնաբերվեցին գլյուկոզա, մաննոզա, ֆրուկտոզա։ Այսպիսով, Ֆիշերի սարտահայտությամբ «գլիցերինը եղավ այն դարպասը, որի միջոցով մենք հասանք բնական շաքարանյութների սինթեզին»։
Ածխածնի մեծ թվով ատոմներ ունեցող մոնոսախարիդները կարելի է ստանալ փոքր թվով ածխածնային ատոմներով ալդոզներից ցիանհիդրինային սինթեզի օգնությամբ։
Բնության մեջ
խմբագրելՄոնոսախարիդները լայնորեն տարածված են բնության մեջ ինչպես ազատ վիճակում, այնպես էլ կապված ձևով։ Առաջնում են պոլիսախարիդների, տանինի տիպի դաբաղանյութերի մեջ, մի շարք բարդ սպիտակուցների՝ նուկլեոպրոտեիդների և գլյուկոպրոտեիդների հիդրոլիզի ժամանակ ստացվում է գլյուկոզա։ Բնության մեջ բացառապես հանդիպում են միայն պենտոզներ և հեքսոզներ։
Ծանոթագրություններ
խմբագրել- ↑ «biochemistry.ru». Արխիվացված է օրիգինալից 2012 թ․ դեկտեմբերի 24-ին. Վերցված է 2015 թ․ մարտի 15-ին.
- ↑ Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. ISBN 0-13-250882-6.
- ↑ Pigman, Ward; Horton, D. (1972). «Chapter 1: Stereochemistry of the Monosaccharides». In Pigman and Horton (ed.). The Carbohydrates: Chemistry and Biochemistry Vol 1A (2nd ed.). San Diego: Academic Press. էջեր 1–67.
Գրականություն
խմբագրել- Օրգանական քիմիայի դասընթաց, Բ. Ա. Պավլով և Ա. Պ.Տերենտև
- Բիոքիմիա, Մ. Գ. Գասպարյան, Ա. Ա. Մնացականյան, Գ. Վ. Բարսեղյան
- McMurry, John. Organic Chemistry. 7th ed. Belmont, CA։ Thomson Brooks/Cole, 2008. Print.
Տես նաև
խմբագրելԱրտաքին հղումներ
խմբագրել| Վիքիպահեստն ունի նյութեր, որոնք վերաբերում են «Մոնոսախարիդներ» հոդվածին։ |